


| name | Amanita sphaerobulbosa |
| name status | nomen acceptum |
| author | Hongo |
| english name | "Asian Abrupt-Bulbed Lepidella" |
| cap | The fruiting bodies of A. sphaerobulbosa are usually small. The cap is 20-40 mm wide, convex to applanate, white to creamy, and with an appendiculate margin. It is covered with conical to subconical, small, white warts that are about 1 mm high; its flesh is white. |
| gills | The gills of this species are free to subfree, crowded, and white to cream-colored; and the short gills are attenuate and of diverse lengths. |
| stem |
The stipe is 50 - 80 × 5 - 8 mm, subcylindric to attenuate upwards; its surface is white to dirty white. The stipe's basal bulb is 15 - 25 mm wide, subabrupt to abrupt, marginate, with the upper margin covered with whitish, small volval remnants. The annulus is membranous and superior, persistent or fugacious. |
| spores |
Spores measure (7.0-) 8.0-10.5 (-11.5) × (6.5-) 7.0-9.5 (-10.0) µm and are globose, subglobose to broadly ellipsoid, and amyloid. Clamps are present on the bases of basidia. |
| discussion |
Amanita sphaerobulbosa was originally described from Japan. It may also occur in China, but no authentic material has been examined. It occurs in mixed forests with broad-leaved trees and conifers. Amanita sphaerobulbosa has been reported as the cause of severe poisoning in Japan. The symptoms suggest the allenic norleucine syndrome known from A. smithiana Bas. Amanita sphaerobulbosa is very similar to A. abrupta Peck, and the names have often been regarded as synonyms. Undoubtedly, they are very closely related to each other. However, the spores of A. abrupta are much narrower. Furthermore, in A. sphaerobulbosa, many hyphae and inflated cells, especially in the upper part of volval remnants on pileus, are vascular (oleiferous), i.e., have refractive contents; this is not the case in A. abrupta. The partial veil of A. abrupta is very persistent (rarely, if ever, fugacioius).—Z. L. Yang |
| brief editors | RET |
| name | Amanita sphaerobulbosa | ||||||||
| author | Hongo. 1969. J. Jap. Bot. 44: 230, fig. 1(1-4). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Asian Abrupt-Bulbed Lepidella" | ||||||||
| MycoBank nos. | 326114 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | TNS [per Doi. 1991. Bull. Natl. Sci. Mus., Tokyo, Ser. B 17(2): 50] | ||||||||
| revisions | Z. L. Yang and Y. Doi. 1999. Bull. Natl. Sci. Mus. Tokyo B 25(3): 119-120, figs. 17-19. | ||||||||
| selected illustrations | Imazeki and Hongo. 1987. Color. Illus. Mushr. Japan 1: 132, pl. 33 (fig. 229). [As "Amanita abrupta."] | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of Dr. Z. L. Yang or another researcher is based upon original research by R. E. Tulloss. NOTE: Spore measurements from papers by Z. L. Yang use his "Times New Roman" face for "Q" and "Q'"—respectively, " | ||||||||
| basidiospores |
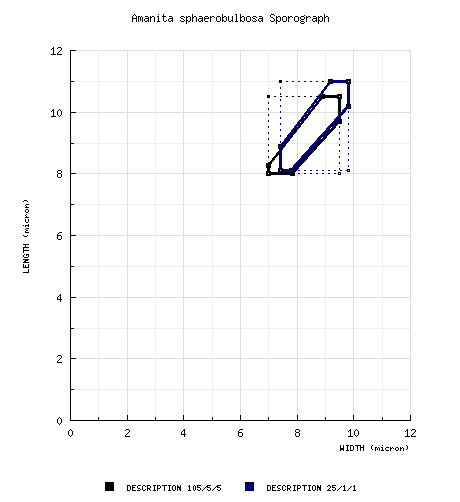
From Yang and Doi (1999): [105/5/5] (7.0-) 8.0 - 10.5 (-11.5) × (6.5-) 7.0 - 9.5 (-10.0) μm, ( composite spore data from material revised by RET: [25/1/1] (7.8-) 8.1 - 11.0 (-11.9) × 7.4 - 9.8 (-10.3) μm, (L = 9.2 μm; W = 8.4 μm; Q = 1.04 - 1.20 (-1.26); Q = 1.10), subhyaline, colorless, smooth, thin-walled, amyloid, globose to subglobose to broadly ellipsoid, often adaxially flattened, with "giant" spores sometimes present; apiculus sublateral, cylindric; contents multiguttulate; color in deposit not recorded. | ||||||||
| ecology | Japan: In Fagus forest or in Quercus forest. | ||||||||
| material examined |
from protolog: JAPAN: HONSHU—Shiga-ken - Mt. Watamuki, 17.ix.1967 T. Hongo 3504 (holotype, in herb. Hongo => TNS F-244377). from Yang and Doi (1999): JAPAN: HONSHU—Shiga-ken - Mt. Watamuki, 17.ix.1967 T. Hongo 3504 (holotype, in herb. Hongo => TNS F-244377). Tokyo Metropolis (Pref.) - Oume-shi, Kurosawa 3-chôme, Nagayama-Kyûryô Hills, along hiking course, 5.x.1997 Y. Doi s.n. (TNS F-237770); Oume-shi, Kurosawa 3-chôme, right side of Kurosawa-gawa R., 25.ix.1997 Y. Doi S.n. (TNS F-237949), 2.viii.1997 Y. Doi & K. Miyazaki s.n. (TNS F-182502); Oume-shi, Kurosawa 3-chôme, Nagayama-Kyûryô Hills, left-side of Kurosawa-gawa R., along hiking course, 5.x.1997 Y. Doi & K. Miyazaki s.n. (TNS F-237569). RET: JAPAN: UNKN. ISL.—Unkn. Prefecture - unkn. loc., s.d. unkn. coll. s.n. [Borovička B 381] (PRM, RET 455-10). | ||||||||
| discussion |
For a number of years this species was confused with the North American species A. abrupta; however, the two taxa are now seen as clearly distinct. They are both assignable to Bas' stirps Microlepis. It would be very interesting to know if a syringaldazine spot text of the east Asian species would produce the dramatic purple (positive) response for the presence of laccase that the North American taxon produces. Sporographs of the two species are compared in the following figure: The Japanese name for the present species is "tama-shiro-onitake" (Doi 1991). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.