


| name | Amanita sp-Kerala01 |
| name status | cryptonomen temporarium |
| author | K. B. Vrinda & Tulloss |
| images |
 1. Amanita sp-Kerala01, rain-bleached specimens, Acacia plantation, Thiruvananthapuram, Peyramam, Kerala, India.  2. Amanita sp-Kerala01, Acacia plantation, Thiruvananthapuram, Peyramam, Kerala, India. |
| intro | The following is derived from the original research of K. B. Vrinda and R. E. Tulloss. |
| cap | The dark gray to grayish beige cap is 35-65 mm wide and streaked with radial fibers. The cap is sometimes white after rain, and in dry weather the cap's skin may split and expose the cap's white flesh. The cap is planar to convex and there is a slight rounded knob in the center of the cap. The white cap's flesh is up to 4 mm thick above the stem. The edge of the cap is not grooved and is decurved in younger specimens and irregular and wavy in older specimens. Volval remnants are sometimes seen as a single large white patch and soon disappear. |
| gills | The crowded gills are free and up to 5 mm broad; they are usually white during all stages of development, but may become yellow with age. The short gills are not cut off squarely and sometimes have an extended narrow tooth-like projections along the underside of the cap. |
| stem | The white stem is 50 - 90 × 4 - 7 mm with brown fibers on its surface below the flaring skirt-like ring; the stem narrows upward slightly. The bulb is egg-shaped and the stem's flesh is white. The three-lobed membranous volva extends up to 30 mm from the bulb base. |
| odor/taste | The odor of this species is not distinctive. The taste has not been recorded. Until demonstrated otherwise, this species should be assumed to contain amatoxins and to be POISONOUS. |
| spores | The spores of this species measure (7.4-) 8.0 - 10.4 (-11.5) × (5.0-) 5.5 - 7.1 (-8.8) μm and are weakly amyloid to amyloid and broadly ellipsoid to ellipsoid (infrequently elongate). There are no clamps at bases of basidia. |
| discussion |
The present taxon was found in groups growing in leaf litter in a plantation of Acacia (imported from Australia) in Kerala India. Morphologically, this species is distinct from all known taxa in sect. Phalloideae. The pattern of coloring on the cap is reminiscent of amatoxin-containing species with streaked caps (e.g., A. phalloides and A. marmorata). Initial genetic sequences from Vrinda S7093 and S9651 match most closely sequences from three collections of A. marmorata.—K. B. Vrinda, R. E. Tulloss, C. Rodríguez Caycedo and N. Goldman |
| brief editors | RET |
| name | Amanita sp-Kerala01 | ||||||||||||||||
| author | K. B. Vrinda & Tulloss | ||||||||||||||||
| name status | cryptonomen temporarium | ||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||
| intro |
Olive text indicates a specimen that has not been
thoroughly examined (for example, for microscopic details) and marks other places in the text
where data is missing or uncertain. The following material is based on Vrinda et al. (2005a) and on original research by the authors of this page. | ||||||||||||||||
| pileus | Vrinda et al. (2005a): 35 - 65 mm wide, gray (4B-C1) to grayish beige (4C2) or greyish brown to yellowish brown (5E3-4), appearing streaked as though with innate radial fibers (although closer examination shows more irregularity in color distribution), sometimes almost white after rain, in dry weather with pileipellis often splitting here and there and exposing white context, convex to planar to depressed in age, with slight umbo, viscid to subviscid when young, becoming dry and shiny when old; context white, up to 4 mm thick above stipe, not bruising or staining on exposure; margin nonappendiculate, nonstriate, regular and decurved in younger specimens, irregular and undulating in older specimens, becoming rimose, sometimes flaring upward in older specimens; universal veil often absent, occasionally as single large white patch and soon disappearing. | ||||||||||||||||
| lamellae |
Vrinda et al. (2005a): free, crowded, white during all stages of development or becoming very pale yellow with age, unchanging when bruised, up to 5 mm broad, ventricose; lamellulae subattenuate or attenuate or attenuate with an extended narrow "tooth" projecting along underside of cap context, moderately common??, of at least four distinct lengths. [Self-reminder to the authors: Need to recharacterize form and quantity of lmaellulae in fresh material.] | ||||||||||||||||
| stipe |
Vrinda et al. (2005a): 50 - 90 × 4 - 7 mm, white, with hair brown (5E4) surface fibrils below partial veil, narrowing upward slightly; bulb ovoid; context white, fibrous, unchanging; partial veil white, membranous, flaring and skirt-like, superior, persistent, striate on upper surface, smooth below, eventually collapsing on stipe; universal veil as membranous limbate volva, white, usually 3-lobed, up to 30 mm from bulb base to highest point on limb. [Self-reminder to the authors: It is desirable to separate the bulb length from the stem length and provide separate measurement for each of the parts.] | ||||||||||||||||
| odor/taste | Vrinda et al. (2005a): Odor not distinctive. Taste not recorded. | ||||||||||||||||
| macrochemical tests |
Vrinda et al. (2005a): H2SO4 on fresh lamellae: purplish tint developing within seconds. KOH - negative on context and surfaces of basidiome. HCL - negative on context and surfaces of basidiome. HNO3 - negative on context and surfaces of basidiome. [Self-reminder to the authors: What were the concentrations of the acids used?] | ||||||||||||||||
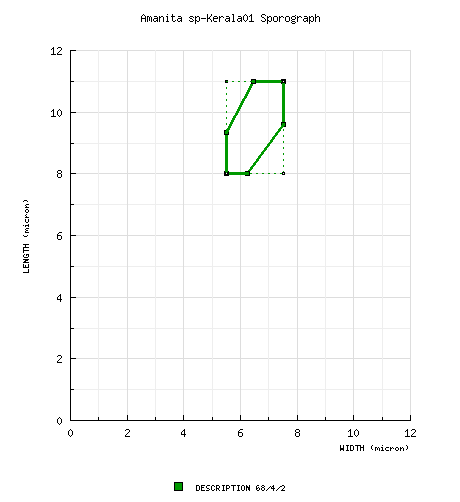
| basidiospores |
Vrinda et al.
(2005a):
[180/9/6] (5.5-) 8.8 - 10.5 (-11.2) ×
(5-) 6.5 - 7.5 (-8.8) μm, (L' = 8.9 μm;
W' = 6.5 μm; Q' = 1.37),
hyaline, thin-walled, smooth, weakly amyloid to
amyloid; white in deposit. [Note: A sporograph
cannot be generated for this data.—ed.]
RET/CRC: [68/4/2] (7.4-) 8.0 - 11.0 (-11.5) × (5.0-) 5.5 - 7.5 (-8.8) μm, (L = 8.8 - 9.7 μm; L' = 9.3 μm; W = 5.9 - 6.4 μm; W' = 6.3 μm; Q = (1.18-) 1.28 - 1.70 (-1.73); Q = 1.47 - 1.51; Q' = 1.48), hyaline, colorless, thin-walled, smooth, weakly amyloid to amyloid, broadly ellipsoid to ellipsoid, infrequently elongate, sometimes swollen at one end, infrequently constricted, Usually adaxially flattened; apiculus sublateral, cylindric; contents granular to guttulate (with or without additional small granules); white in deposit. | ||||||||||||||||
| ecology | Vrinda et al. (2005a): In troops. At ?? m elev. In leaf litter in plantation of Acacia (alien, imported from Australia). | ||||||||||||||||
| material examined |
Vrinda et al. (2005a): INDIA:
KERALA—Perayam - Thiruvananthapuram,
19.iv.2004 K. B. Vrinda S7086 (TBGT),
20.iv.2004 K. B. Vrinda S7087 (TBGT),
21.iv.2004 K. B. Vrinda S7093 (TBGT),
22.iv.2004 K. B. Vrinda S7097 (TBGT),
28.iv.2004 K. B. Vrinda s.n. (TBGT),
29.iv.2004 K. B. Vrinda s7107 (TBGT).
RET: INDIA: KERALA—Perayam - Thiruvananthapuram, 21.ix.2004 K. B. Vrinda S7093 (TBGT; RET 391-7), 2006 K. B. Vrinda S9651 (TBGT; RET 391-9). | ||||||||||||||||
| discussion |
Morphologically, this species is distinct from all
known taxa in sect. Phalloideae.
The pattern of pigmentation on the cap is strongly
reminiscent of amatoxin-containing species with
apparently virgate pilei (e.g., A. phalloides, A.
marmorata, and A. subjunquillea). In the following figures sporographs for the A. phalloides, A. marmorata, and A. subjunquillea are compared with the sporograph of the present species: MegaBLAST runs of partial and complete nrITS sequences from Vrinda S7093 and 9651 produced sequences from three collections of A. marmorata (two from introduced genets in South Africa and one from an introduced genet in Hawaii, U.S.A.) as closest matches, with E value of 0.0 and match percentage of 99%. | ||||||||||||||||
| citations | —K. B. Vrinda, R. E. Tulloss, and C. Rodríguez Caycedo | ||||||||||||||||
| editors | RET | ||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.