


| name | Amanita fulvopulverulenta |
| name status | nomen acceptum |
| author | Beeli |
| english name | "Fulvous-powdered Lepidella" |
| intro | The following description is based on Beeli (1935). |
| cap | The cap of Amanita fulvopulverulenta is 90 - 100 mm wide, flattened-convex, fleshy, with a slightly prominent umbo, with a slightly striate margin. The cap is pale pinkish brown covered with brownish-red, powdery remains of the volva |
| gills | The gills are free, slightly yellowish, and somewhat rounded at the end near the stem. |
| stem | Its stem is 120 - 170 × 10 - 20 mm, solid, and cylindric. The surface is fibrillose to glabrous, white or slightly tinted red. The flesh is whitish but turns pink when cut. The stem is easily separated from the cap. The stem's bulb is spindle-shaped or somewhat turnip or carrot shaped. The limb of volva on the top of the bulb is not very persistent. On the other hand, the ring on the stem is said to be persistent. This is an odd combination of characters for a species that has been thought to be assignable to sect. Amidella. |
| spores | Beeli reported the spores as measuring 7 - 8 × 4 µm, amyloid, and elongate to cylindric. Gilbert's spore measurements are 7.5 - 11.3 × 3.7 - 5.0 µm. |
| discussion |
The present species was originally described from the Democratic
Republic of Congo in dry forests. E.-J. Gilbert's comment that this species is very similar to A. goossensiae Beeli is not borne out by his own spore data (1941). Moreover, as noted above the presence of a bulb and a weak-limbed volva is unusual in sect. Amidella, while the stipe base in A. goossensiae lacks a bulb and is enclosed in a very robust volval sac—characters typical of sect. Amidella On the other hand, the pigments, spore shape, flesh changing to pink when cut, membranous volva that has a powdery inner-layer, and narrow spores are all indicators that have been taken to indicate that a species belongs in sect. Amidella [typified by the American species A. volvata (Peck) Lloyd]. Madame Goossens' painting shows a section of a button with a very distinct bulbous base. In the somewhat macroscopically similar A. goossensiae Beeli and A. fulvosquamulosa Beeli, Madame Goossens carefully distinguished the flesh of the totally elongating stem from the strongly thickened volval sac. That she failed to do so in the case of the present species suggests that it had a true bulb. It should be noted that I have never revised dried material. If Madame Goossens' painting is correct, then A. fulvopulverulenta would be an exception to the dominant stipe and volva development in section Amidella.—R. E. Tulloss, K. Mighell |
| brief editors | RET |
| name | Amanita fulvopulverulenta | ||||||||||||||||
| author | Beeli. 1931. Bull. Soc. Roy. Bot. Belgique 63: 104, pl. 7 (fig. 3). | ||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||
| english name | "Fulvous-powdered Lepidella" | ||||||||||||||||
| synonyms |
≡Amidella fulvopulverulenta (Beeli) E.-J. Gilbert.
1940. Iconogr. Mycol. (Milan 27, suppl. (1): 77,
tab. 28 (fig. 5). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||
| etymology |
fulvus, a reddish, orange- or yellow-brown +
pulverulentus,
covered with powder; hence, covered with fulvous powder In Roman times, fulvus was used for the color of a lion's mane according to some authorities. | ||||||||||||||||
| MycoBank nos. | 212892, 284131 | ||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||
| holotypes | BR (implicit) | ||||||||||||||||
| epitypes | YA [isoepitypes: HSC G1300 and BR 5020224998797V] | ||||||||||||||||
| epitypifications | Mighell et al. 2020. Mycologia 11 Nov. 2020: 7. | ||||||||||||||||
| type studies | Mighell et al. 2020. Mycologia 11 Nov. 2020: 7. | ||||||||||||||||
| selected illustrations | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl.: 311, tab. 38. | ||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present taxon, (Beeli 1935), and (Gilbert 1940 & 1941). | ||||||||||||||||
| pileus |
protolog: 90 - 100 mm wide,
pale flesh-colored, plano-convex; context
fibrous-fleshy, white; margin lightly striate;
universal veil as brownish red pulverulence covering
surface. Mighell et al. (2020): epitype: pale tan-brown (4A-B3) with pinkish tint or brick red (8D8), bruising dark red, 45 - 75 mm across opening and 10 - 34 mm "tall," dry to moist; margin entire, decurved to uplifted, appendiculate with appendiculae 1 - 5 mm long, red-brown, occasionally white tipped [possibly enitrely white at first?—ed.], in pendant palisade coalescing into triangular or rectangular flaps 3 - 8 mm wide; context 0.5 mm thick at margin and 5 mm thick above stipe, solid, pale pinkish white to red; universal veil pulverulent, detersile, as light brown (5B-C3) to dark orange-brown (5-6E8) patches, 0.5 - 2 mm wide, often lost over disc, rarely erect and blunt over disc, nearly continuous and pulverulent near margin. | ||||||||||||||||
| lamellae |
protolog: free, density not described,
lightly yellowish, narrowing toward the stipe, 8 - 9 mm
broad; lamellulae not described. Mighell et al. (2020): epitype: adnexed, crowded, light yellow (4A2), up to 5 mm broad, 1.5 mm broad at ends, with edges white to concolorous and minutely fimbriate; lamellulae attenuate, 1 - 31 mm long. | ||||||||||||||||
| stipe |
protolog:
120 - 170 × 10 - 20 mm, white or
tinged reddish, cylindric, fibrous, glabrous, easily
detachable from pileus; bulb ventricose or
narrowly napiform with radicating base (per figure);
context fibrous-fleshy, white, becoming pinkish
when sectioned; partial veil membranous, white,
thin; universal veil as limbate volva (per figure),
membranous, thin, brown-red, not very
persistent. [Note: A thin, ephemeral, limbate volva on a species with a persistent partial veil and a distinctly bulbous stipe base is quite atypical of Amanita sect. Amidella.—ed.] Mighell et al. (2020): epitype: 66 - 210 × 6.5 - 10 mm, off-white to occasionally pinkish (7-8A2) over upper half, pink (8A5) to brick red (8C-D8) below, cylindric, with appressed fibrils on surface becoming denser and floccose toward bulb and bruising medium brick red; context ...; partial veil ...; bulb 35 - 60 × 15 - 25 mm, subglobose to ovoid to fusiform, with rounded tip, slightly dog-legged with respect to vertical stipe [per images and correspondence&emdash;ed.]; universal veil enclosing bulb, with limb(s) arising near bulb apex and partially or completely enclosing stipe base, occasionally encircled by subconcentric rings of deep pink-red (9B7) recurved floccose squamules, glabrous below and subtended by white hyphal cords. | ||||||||||||||||
| columnella | double click in markup mode to edit. | ||||||||||||||||
| odor/taste |
protolog: Odor strongly
acrid. Taste bitter. Mighell et al. (2020): epitype: of chlorine, notable at a distance of several meters. | ||||||||||||||||
| macrochemical tests |
none recorded. | ||||||||||||||||
| pileipellis | Beeli (1935): filamentous hyphae, thin; inflated cells terminal. [Note: It is possible that inflated cells of the volva or acrophysalides from the pileus context were perceived as included in the pileipellis.—ed.] | ||||||||||||||||
| pileus context | not described. | ||||||||||||||||
| lamella trama | not described. | ||||||||||||||||
| subhymenium | not described. | ||||||||||||||||
| basidia | not described. | ||||||||||||||||
| universal veil | not described. | ||||||||||||||||
| stipe context | not described. | ||||||||||||||||
| partial veil | not described. | ||||||||||||||||
| lamella edge tissue |
protolog: not described. Mighell et al. (2020): | ||||||||||||||||
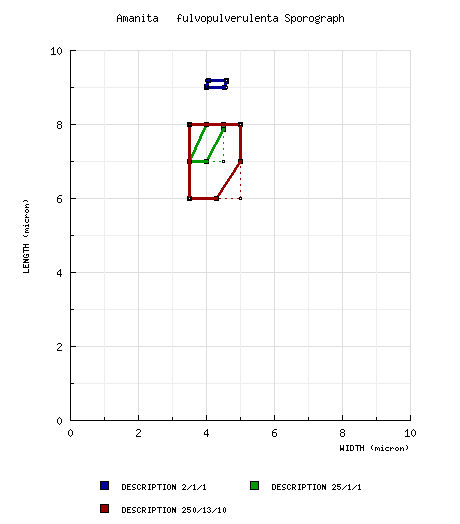
| basidiospores |
from protolog: 7 - 8 × 4 μm,
hyaline, smooth, amyloid. [Note: Beeli's spore data
often appears to be incorrect (values too low).
Compare with values derived from measuring the scaled
drawings of spores in (Gilbert
1940),
below. Sporograph not generated.—ed.] Beeli (1935): differs from the above by saying the spores were inamyloid—apparently an error. Gilbert (1940 & 1941): [2/1/1] 9.0 - 9.2 (-11.5) × 4.0 - 4.6 (-5.1) μm, (Q = 1.99- 2.26), hyaline, smooth, "regular," amyloid, elongate to cylindric, at least somewhat adaxially flattened (per figure); apiculus sublateral (per figure); contents not described; white in deposit. Mighell et al. (2020) from holotype: [25/1/1] (6.5-) 7 - 8 × 3.5 - 4.5 μm, (Q = 1.75 - 2.0; Q' = 1.89). Mighell et al. (2020) from epitype: [250/13/10] 6 - 8 (-9) × (3-) 3.5 - 5 μm, (Q = 1.40 - 2.29; Q' = 1.78), hyaline, smooth, thin-walled, amyloid, pronouncedly flattened adaxially; apiculus 1 μm long; color in deposit probably white. | ||||||||||||||||
| ecology |
from protolog: Dem. Rep. Congo:
Scattered. On soil in dry forest. Mighell et al. (2019): Cameroon: Solitary to scattered on upper mineral soil in forest dominated by Gilbertiodendron dewevrei. | ||||||||||||||||
| material examined |
protolog: DEM. REP.
CONGO: MONGALA—Binga [2.3947° N/
20.4236° E, 361 m],
Beeli (1935): DEM. REP. CONGO: MONGALA—Binga [2.3947° N/ 20.4236° E, 361 m], Gilbert (1940 & 1941): DEM. REP. CONGO: MONGALA—Binga [2.3947° N/ 20.4236° E, 361 m], Mighell et al. (2019): CAMEROON: EAST REGION—Dja Biosphere Reserve - Northwest Sector ca. Somalomo, Upper Dja River Basin, within 2 km of base camp at [3.3560° N/ 12.7297° W, 650 m], 26.viii.2014 Henkel 9887 (epitype, YA; isoepitype HSC G1300; isoepitype BR 5020224998797V). DEM. REP. CONGO: MONGALA—Binga [2.3947° N/ 20.4236° E, 361 m], | ||||||||||||||||
| discussion |
"The powdery inner layer of the universal veil, the
pigments, the color change of the context, the striate
pileus margin, the narrow amyloid spores all appear to
argue for placement of the present taxon in sect.
Amidella. The stipe's having a distinct,
radicating bulb with a fragile, limbate volva encircling
the stipe base and a persistent partial veil could
support placement of the present species in
Amanita
[sect. Lepidella] subsect. Limbatulae,
in which it would be the first taxon with a
pulverulent volval layer that is distinctly
pigmented. "The present species raises the interesting possibility that it could be assignable to subsect. Limbatulae and be basal to sect. Amidella and represent a relict transitional form between the two groups. At any rate, it is very clear that, contrary to some past views, the present species is quite distinct from A. goossensiae and A. fulvosquamulosa." For historical purposes, I have left the above paragraphs unchanged after the proposal of a well-founded epitype for the present taxon. The epitype and molecular research on it strongly support the reclassification proposed here. And the reclassification has now been reflected in the present page and tab. | ||||||||||||||||
| citations | —R. E. Tulloss and Kennan Mighell | ||||||||||||||||
| editors | RET | ||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita fulvopulverulenta |
| bottom links | [ Keys & Checklists ] |
| name | Amanita fulvopulverulenta |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.