


| name | Amanita castanopsis | ||||||||
| author | ("castanopsidis") Hongo. 1974a. Bull. Soc. Linn. Bordeaux, No. Spéc., Trav. Mycol. Dédiés Kühner: 192, fig. 2(A-C). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Asian Chinkapin Lepidella" | ||||||||
| MycoBank nos. | 308544 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | TNS [per Doi. 1991. Bull. Natl. Sci. Mus., Tokyo, Ser. B 17(2): 49] | ||||||||
| type studies | Z. L. Yang and Y. Doi. 1999. Bull. Natl. Sci. Mus. Tokyo B 25(3): 117. | ||||||||
| revisions | Z. L. Yang. 1997. Biblioth. Mycol. 170: 142-143. | ||||||||
| selected illustrations | Imazeki and Hongo. 1987. Color. Illus. Mushr. Japan 1: 134, pl. 34 (fig. 233). | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present taxon, the type study of Yang and Doi (1999), (Yang 1997), and (Yang 2005). NOTE: Spore measurements from papers by Z. L. Yang use his "Times New Roman" face for "Q" and "Q'"—respectively, " | ||||||||
| pileus |
from protolog: 35 - 75 mm wide, pure white, convex, then planar, dry; context white, "moderately thick"; margin nonstriate, appendiculate; universal veil as warts, acute, conic or pyramidal, white with tips often becoming grayish or brownish, densely set, adnate, floccose, 1 - 3 × 1 - 4 mm, densely set, decreasing in size toward margin. from Yang (1997): ca. 80 mm wide, white, convex to subplanar; context white, becoming thinner toward margin; margin nonstriate, appendiculate; universal veil as abundant white adnate warts, subglobose to pyramidal, 1 - 3 × 1 -4 mm, floccose, becoming smaller toward margin. | ||||||||
| lamellae |
from protolog: free with decurrent line, subdistant, white, 6± broad, with white flocculose edge; lamellulae not described. from Yang (1997): free to nearly free, not crowded, white, 4 - 6 mm broad, narrowing toward stipe; lamellulae 0 - 3 between pair of otherwise adjacent lamellae, with shorter ones truncate (but not squarely), with longer ones attenuate to subattenuate. | ||||||||
| stipe |
from protolog: 70 - 80 × 10 - 15 mm (width measured at apex), white, floccose-pruinose; bulb clavate or broadly ventricose with pointed base, 25 - 35 mm wide; context white, rather firm, solid or stuffed; partial veil thick, white, "floccose-cobwebby, friable, mostly disappearing on expansion of pileus"; universal veil as more or less concentric rings of small floccose granules or subconical warts. from Yang (1997): 80± × 10 - 15 μm, white, subcylindric, robust; bulb napiform to ventricose, 20 - 30 μm wide; context white; partial veil soon in tatters; universal veil as white subconic warts girdling upper half of stipe's bulb. | ||||||||
| odor/taste |
from protolog: Odor and taste lacking. from Yang (1997): Odor and taste inconspicuous. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | from Yang (1997): poorly differentiated, gradually merging with pileus context; filamentous hyphae 2 - 6 μm wide, disordered, interwoven, somewhat gelatinized; vascular hyphae rare. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | from Yang (1997): more or less bilateral; with central stratum comprising filamentous hyphae (2 - 7 μm wide, interwoven, frequently branching, rather abundant to abundant) and inflated cells [rather abundant to abundant, narrowly ventricose to broadly clavate, 50 -90 × 16 - 20 μm, intercalary or (rarely) terminal] and vascular hyphae (4 - 10 μm wide, rare or rather abundant in scattered localities); with lateral strata comprising filamentous hyphae (3 - 6 μm wide, rather abundant) and inflated cells (clavate to narrowly ventricose, 45 - 60 × 12 - 17 μm); clamps common. | ||||||||
| subhymenium | from Yang (1997): not well delimited; inflated cells 15 -35 × 12 - 16 μm, ellipsoid to pyriform or irregularly shaped, with scattered intermixed partially inflated cells 5 - 8 μm wide. | ||||||||
| basidia |
from protolog: 42 - 53 × 9.5 - 10 μm, 4-sterigmate; clamps present. from Yang and Doi (1999): 40 - 55 × 9.0 - 11.0 μm, 4-sterigmate, with sterigmata 3.0 - 5.0 μm long; clamps present. from Yang (1997): 45 - 56 × 11 - 14 μm, predominately 4-, rarely 1- or 2-sterigmate, with sterigmata 4 - 5 μm long; clamps frequent. | ||||||||
| universal veil |
from protolog: On pileus: filamentous hyphae 3.6 - 7.7 μm wide, branching; inflated cells 20 - 70 × 15 - 43 μm, clavate to ellipsoid to subglobose, terminal singly or in short rows. from Yang and Doi (1999): On pileus: elements anticlinally aligned; filamentous hyphae 3 - 5 μm wide, locally fairly abundant, thin-walled, colorless, hyaline; inflated cells abundant to dominant, subglobose to ovoid to ellipsoid, 35 - 75 × 25 - 50 μm, terminal singly or in chains of 2 - 3 (-4), thin-walled, colorless, hyaline to subhyaline; vascular hyphae rare; clamps often present. On stipe: not recorded. from Yang (1997): On pileus: elements anticlinally aligned; filamentous hyphae 2 - 5 μm wide, abundant, loosely organized, frequently branching; inflated cells abundant, subglobose to ovoid, (30 - 65 × 28 - 53 μm) to ellipsoid (50 - 72 × 24 - 47 μm) to elongate-ellipsoid or broadly clavate (60 - 95 × 32 - 41 μm), often in short chains; vascular hyphae rare. On stipe: not recorded. | ||||||||
| stipe context | from Yang (1997): longitudinally acrophysalidic; filamentous hyphae 2 - 7 μm wide, branching; acrophysalides 200 - 250 × 24 - 30 μm, abundant; vascular hyphae rare or solitary and irregularly sinuous; clamps not abundant. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | not described. | ||||||||
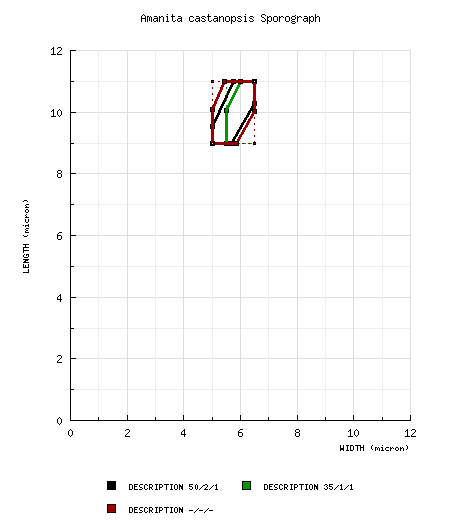
| basidiospores |
from Yang and Doi (1999): [50/2/1] (8.5-) 9.0 - 11.0 × 5.0 - 6.5 μm, ( from Yang (1997): [35/1/1] (8.5-) 9.0 - 11.0 (-12.0) × (5.0-) 5.5 - 6.5 μm, ( from (Yang 2005): [-/-/-] (8.5-) 9.0 - 11.0 (-12.5) × (4.5-) 5.0 - 6.5 μm, ( | ||||||||
| ecology |
from protolog: Japan: On bare red clay in forest of Castanopsis cuspidata. from Yang (1997): China: At 2500± m elev. In broad-leaved forest. | ||||||||
| material examined |
from Yang and Doi (1999): JAPAN: HONSHU—Ôtsu, Kokubu, 6.viii.1970 T. Hongo 4198 (paratype, in herb. T. Hongo), 7.viii.1971 T. Hongo 4493 (paratype, in herb. T. Hongo), 18.ix.1972 T. Hongo 4753 (holotype, in herb. T. Hongo => TNS F-237275). from Yang (1997): CHINA: YUNNAN—Dali Bai Autonomous Prefecture - Binshuan Co., Jizushan, 2500 m elev., 9.viii.1985 G. P. Xiao 521 (HKAS 17037). | ||||||||
| discussion |
The anticlinally organized elements of the universal veil on the pileus in this species segregates it from the taxa of Bas' stirps Virgineoides to which there is otherwise some similarity as noted by Yang (1997) and Yang and Doi (1999). Among the stirpes proposed by Bas, the one to which the present taxon seems most likely to belong is stirps Microlepis. more t.b.d. The Japanese name for this species is "ko-shiro-onitake" (Doi 1991). | ||||||||
| citations | Translation from German by RET.—Zhu L. Yang and R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita castanopsis |
| bottom links | [ Keys & Checklists ] |
| name | Amanita castanopsis |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.
