


| name | Amanita russuloides |
| name status | nomen acceptum |
| author | (Peck) Sacc. |
| english name | "Russula-like Amanita" |
| intro | The following is based on a description by Jenkins included in his type study of A. russuloides (Jenkins 1978a). At present, RET has probably collected material that could resolve the identity of this species, but has not had time to process it; hence, a modern description of the taxon is lacking. |
| cap | The cap of A. russuloides is approximately 25 mm wide, pale yellow to straw yellow, convex to plano-convex, with a striate margin. The volva is present as a few widely scattered remnants. |
| gills | The white gills are crowded, free, but connected to the stem by a floccose line. The short gills are truncate. |
| stem | The stem is approximately 35 × 2 - 5 mm, slightly tapering upward, stuffed, smooth, with a bulbous base. No ring was present in the material reviewed by Jenkins. The volva is present as a slight free limb at the top of the bulb and as occasional floccose patches on the lower stem. |
| odor/taste | double click in markup mode to edit. |
| spores | According to Jenkins' type study, the spores measure 8.7 - 10.2 × 6.3 - 7.0 µm and are broadly ellipsoid to ellipsoid and inamyloid. |
| discussion |
This species was originally described from New York (state), U.S.A. The above, limited description has made it quite difficult to develop a species concept that is applicable to the present name. It has also proven difficult (to date) to sort through the yellow taxa of eastern North America with a view to identifying a single, most likely candidate for the name "A. russuloides." RET is not even sure how many such taxa there are. Moreover, insufficient time has been spent on this problem. Of RET's numbered taxa listed in checklists and keys, there are at least three incompletely understood candidates: A. sp-N08, A. sp-N19, and A. sp-S01. All three of these taxa have spores that are predominantly ellipsoid, with some broadly ellipsoid and some elongate; and, in two, cylindric spores have been observed rarely. All three have macroscopic descriptions that are reasonable matches for A. russulolides. Do all the specimens of the three "numbered taxa" correspond to a single species? The spore data for A. sp-N19 and A. sp-S01 are very similar. Do one or more of the provisional, numbered species correspond to A. russuloides? The questions are as yet unanswered.—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita russuloides | ||||||||
| author | (Peck) Sacc. 1887. Syll. Fung. 5: 13. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Russula-like Amanita" | ||||||||
| synonyms |
≡Agaricus russuloides Peck. 1873 (April) ("1874"). Bull. Buffalo Soc. Nat. Sci. 1(2): 41. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 172466 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | NYS | ||||||||
| type studies |
Jenkins. 1977. Biblioth. Mycol 57: 97. [Jenkins considered this name a synonym of Amanita gemmata (Fr.) Bertillon in DeChambre.] Jenkins. 1978a. Mycotaxon 7: 40. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present taxon, (Jenkins 1977), and (Jenkins 1978a). Any other material not cited as the work of another researcher is based on original research of R. E. Tulloss. | ||||||||
| pileus | type study of Jenkins (1978a): ca. 35 mm wide, pale yellow to straw color, convex to plano-convex; context ?; margin striate; universal veil as widely scattered floccose patches. | ||||||||
| lamellae | type study of Jenkins (1978a): free but connected to stipe by floccose line, crowded, white; lamellulae truncate. | ||||||||
| stipe | type study of Jenkins (1978a): ca. 35 × 2 - 5 mm, tapering slightly upward, smooth; bulb present; context stuffed; exannulate; universal veil as slight free limb at bulb apex, with occasional patches on lower stipe. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
spot test for laccase (syringaldazine) - negative in all parts of basidiome at both intermediate and mature stages of development; spot test for tyrosinace (paracresol) - rapidly positive throughout most of basidiome with exception of lamellae (eventually positive) and universal veil (negative or weakly positive) in basidiomes at both intermediate and mature stages of development. Test voucher: Tulloss 9-9-99-F. | ||||||||
| pileipellis | type study of Jenkins (1977, 1978a): filamentous hyphae densely interwoven to subradial, gelatinized. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | type study of Jenkins (1977, 1978a): bilateral; filamentous hyphae 3 - 8 μm, moderately branched; inflated cells up to 130 × 25 μm, mostly clavate to irregularly elongate; clamps not observed. | ||||||||
| subhymenium | type study of Jenkins (1977, 1978a): ramose; clamps not observed. | ||||||||
| basidia | type study of Jenkins (1977, 1978a): 39 - 50 × 4.1 - 12 μm, 4-sterigmate; clamps rare. | ||||||||
| universal veil | type study of Jenkins (1977, 1978a): On pileus: loose to fairly dense tissue, with elements anticlinally oriented to disordered; filamentous hyphae 4 - 8 μm, moderately branched; inflated cells clavate to ellipsoid to oblong-ellipsoid to astringo-cylindric (up to 110 × 32 μm), broadly ellipsoid to ovoid (up to 64 × 40 μm), terminal in chains; clamps not observed. On stipe base: very similar to tissue on pileus; filamentous hyphae 3 - 8 μm wide, moderately branched; inflated cella as on pileus but disposed irregularly and with ellipsoid and ovoid cells larger; clamps rare. | ||||||||
| stipe context | type study of Jenkins (1977, 1978a): longitudinally acrophysalidic; filamentous hyphae 3 - 8 μm wide, moderately branched; acrophysalides up to 255 × 32 μm; clamps rare. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue | not described. | ||||||||
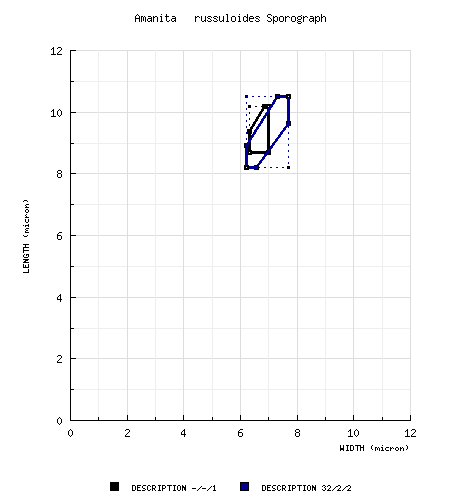
| basidiospores |
type study of Jenkins (1978a): [-/-/1] 8.7 - 10.2 × 6.3 - 7.0 μm, (Q = 1.24 - 1.49; Q' = 1.40), hyaline, smooth, thin-walled, nonamyloid, broadly ellipsoid to ellipsoid, adaxially flattened; apiculus sublateral, cylindric; contents guttulate; color in deposit not recorded. composite of data from all material revised by RET: [32/2/2] (8.0-) 8.2 - 10.5 (-10.8) × (6.0-) 6.2 - 7.7 (-7.7) μm, (L = 8.8 - 10.0 μm; L' = 9.2 μm; W = 6.7 - 7.2 μm; W' = 6.8 μm; Q = (1.23-) 1.25 - 1.44 (-1.53); Q = 1.32 - 1.40; Q' = 1.35), hyaline, colorless, smooth, thin-walled, inamyloid, broadly ellipsoid to ellipsoid; apiculus sublateral, ??; contents ??; ?? in deposit. | ||||||||
| ecology | not recorded. | ||||||||
| material examined |
from type study of Jenkins (1978a):
U. S. A.: NEW YORK— Rensselaer Co. - Greenbush,
s.d. C. H. Peck s.n. (holotype, NYS). RET: U.S.A.: CONNECTICUT—New Haven Co. - Cheshire Twp., Sleeping Giant St. Pk. [41°26'02" N/ 72°53'03" W, 179 m], 23.vii.1992 NEMF1992 participant s.n. [Tulloss 7-23-92-B] (RET 066-5) (RET). NEW JERSEY—Monmouth Co. - Upper Freehold Twp., Assunpink Wildlife Management Area [40°12’36” N/ 74°28’42” W, 53 m], 9.ix.1999 M. A. & R. E. Tulloss 9-9-99-F (RET 299-3). Salem Co. - unkn. loc., 29.vii.1994 Mr. & Mrs. Masi s.n. [Tulloss 7-29-94-A] (RET). NEW YORK—Essex Co. - Keene, 21.viii.1987 Nuckel s.n. [Tulloss 8-21-87-M] (RET). | ||||||||
| discussion |
Jenkins (1977): "The specimen citation in the original description is very incomplete and under ordinary circumstances would not suffice as a citation. However, this taxon appears to be rare and Peck (1906) later stated that he had made no additional collections at Greenbush. Under these conditions I feel that the citation justifies the matching packet at NYS as a holotype (Lanjouw, 1966: Guide for the Determination of Types)." The material that RET assigned here (see the materials examined data field, above) are collections with spore measurements most like that reported by Jenkins in his type study of A. russuloides. This is probably an overly restrictive view of the spores of the present species. The holotype of A. russuloides was probably collected in the early 1870's. Spores from an old type collection may often be found to be smaller than spores in recently collected material of the same species. Perhaps, Amanita sp-N08, A. sp-N19, and/or A. sp-S01 belong here. In the following figures, the known spore data from the numbered provisional taxa are compared with the data provided in the basidiospores data field on the present page: Of the "gemmatoid taxa" listed on this site, one North American species with spore data particularly close to the data on this tab is A. sp-T22—a small species that has had a beige rather than yellow pileus in the two collections seen by RET. Because of the very small basidiomes, pileus pigmentation, and apparent lack of a volval limb on the basal bulb of the stipe, A. sp-T22 is currently segregated from the material provisionally included in A. russuloides. In addition to dealing with the (possibly too large) number of code-numbered, provisional, gemmatoid taxa, gaining an undertsanding for the present species may also require a thorough study of material that might be assigned to another North American taxon—A. agglutinata. As noted on the brief tab of the present page, resolution of the nature of A. russuloides is dependent (at minimum) on future anatomical studies. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita russuloides |
| bottom links | [ Keys & Checklists ] |
| name | Amanita russuloides |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.