


| name | Amanita flavipes |
| name status | nomen acceptum |
| author | S. Imai |
| english name | "Asian Yellow Dust Amanita" |
| images |
 1. Amanita flavipes, Yunnan Prov., China. |
| cap |
The fruiting bodies of A. flavipes are small to medium-sized. The cap is 30 - 120 mm wide, convex to applanate, yellowish to yellow to yellowish brown to brownish orange, darker in the center, paler towards the margin, sometimes with grayish tints. It is covered with yellowish to yellow to yellowish white, occasionally whitish, floccose, felty, granular volval warts; its context is white to whitish. |
| gills |
The gills of this species are free to subfree, crowded, white to yellowish; and the short gills are attenuate and of diverse lengths. |
| stem |
The stipe is 40 - 150 × 5 - 20 mm, subcylindrical to attenuate upwards; its surface is white to yellowish to yellow, covered with yellowish to yellow squamules, and glabrescent; the stipe's basal bulb is 15 - 40 mm wide, subglobose to ovoid to ventricose, with the upper part covered with yellowish, farinose to granular volval remnants in a few concentric belts. The annulus is membranous and superior with its upper surface yellow to yellowish to cream and lower surface yellowish to yellow. The species occurs in conifer forest or in forests with oaks and conifers. |
| spores |
Spores [East Asian] measure (6.5) 7.0 - 9.0 (10.0) × (5.0) 5.5 - 7.0 (8.5) µm and are broadly ellipsoid to ellipsoid and amyloid. [South Asian spore measurements are as follows: (6.0-) 6.8 - 8.5 (-11.0) × (5.0-) 5.2 - 6.5 (-8.8) µm.] Clamps are not present on the bases of basidia. |
| discussion |
Amanita flavipes was originally described from Japan. The species is widely distributed in China, India, Japan, Pakistan, and South Korea. For comparison, see A. elongata Peck, Amanita erythrocephala Neville, Poumarat & Aste, A. flavoconia G. F. Atk., and A. flavoconia var. inquinata Tulloss, Ovrebo & Halling. L. F. Zhang, J. B Yang and Zhu L. Yang (2004) recently published molecular evidence that Amanita flavipes comprises a number of genetic clades that have macroscopic differences such as pigmentation as well as differences first detected on the molecular level. The reader may wish to reference the discussion following the description of A. strobilaceoides A. E. Wood.—Zhu L. Yang and R. E. Tulloss |
| brief editors | RET |
| name | Amanita flavipes | ||||||||
| author | S. Imai. 1933. Bot. Mag. (Tokyo) 47: 428. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Asian Yellow Dust Amanita" | ||||||||
| synonyms |
≡Amplariella flavipes (S. Imai) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 79, tab. 47 (fig. 3).
=Amanita bella S. Imai in E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 47, tab. 47(fig. 4). ≡Amplariella bella (S. Imai in E.-J. Gilbert) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 79. ≡Amanita pulchella S. Imai nom. illeg 1933. Bot. Mag. (Tokyo) 47: 427. non
Amanita pulchella (Cooke & Massee) E.-J. Gilbert (=Amanita xanthocephala)
=Amanita watlingii A. Kumar & T. N. Lakh. in A. Kumar et al. 1990. Amanitaceae India: 92, 112, fig. 18(A-E).
=Amanita flavoconia sensu A. Kumar et al. 1990. ibid.: 63, 99, fig. 5(A-E).
=Amanita flavorubescens sensu Yuan & Sun. 1995. Sichuan Mushrooms: 495. non Amanita flavorubescens G. F. Atk. (=A. flavorubens) The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 265044, 284153 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
A. bella—in herb. S. Imai (SAP); isotype (fragment) in herb. Gilbert (lost). A. watlingii—HPUB. | ||||||||
| lectotypes | A. flavipes—in herb. S. Imai (SAP); isolectotype (fragment), in herb. Gilbert (lost). | ||||||||
| lectotypifications | E.-J. Gilbert. 1940. op. cit.: 174 (in caption to fig. 3). | ||||||||
| revisions |
Z. L. Yang. 1997. Biblioth. Mycol. 170: 202, figs. 169-172. Tulloss et al. 2001. Mycotaxon 77: 480, figs. 13-15. | ||||||||
| selected illustrations |
E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl.: tab. 54-55. Imazeki and Hongo. 1965. Color Illus. Fungi 2: 41, pl. 12(fig. 75). Imazeki et al. 1988. Fungi Japan: 162. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. Any of the following material not directly from the protolog of the present taxon and not cited as the published work of Dr. Z. L. Yang or another researcher is based upon original research by R. E. Tulloss. NOTE: Spore measurements from papers by Z. L. Yang use his "Times New Roman" face for "Q" and "Q'"—respectively, " | ||||||||
| basidiospores |
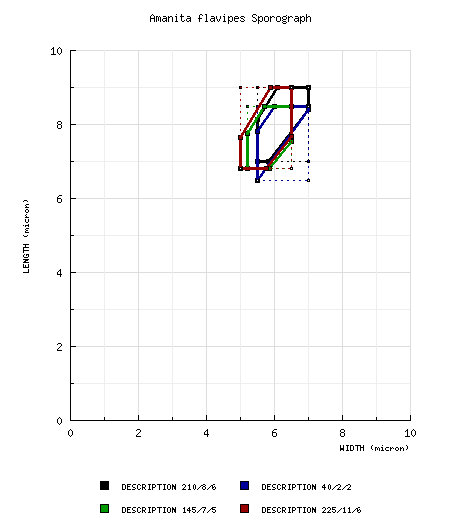
from Yang (1997): [210/8/6] (6.5-) 7.0 - 9.0 (-10.0) × (5.0-) 5.5 - 7.0 (-8.5) μm, ( from Yang (2002a): [40/2/2] 6.5 - 8.5 (-9.0) × (5.0-) 5.5 - 7.0 μm, ( from Tulloss et al. (2001): [145/7/5] (6.0-) 6.8 - 8.5 (-11.0) × (5.0-) 5.2 - 6.5 (-8.8) µm, (L = 7.3 - 7.7 (-7.8) µm; L’ = 7.5 µm; W = 5.7 - 5.9 µm; W’ = 5.8 µm; Q = (1.12-) 1.16 - 1.49 (-1.69); Q = 1.27 - 1.34 (-1.38); Q’ = 1.30), hyaline, colorless, thin-walled, smooth, amyloid, broadly ellipsoid to ellipsoid, occasionally subglobose or elongate, often adaxially flattened, sometimes swollen at one end; apiculus sublateral, cylindric, narrow, sometimes prominent; contents mono- or multiguttulate, with or without additional small granules; white in deposit. Composite of data from India and Pakistan (RET): [225/11/6] (5.8-) 6.8 - 9.0 (-11.0) × (4.2-) 5.0 - 6.5 (-8.8) μm, (L = 7.3 - 7.9 (-8.2) μm; L' = 7.6 μm; W = (5.4-) 5.6 - 5.9 μm; W' = 5.7 μm; Q = (1.12-) 1.18 - 1.53 (-1.73); Q = 1.27 - 1.40; Q' = 1.33), colorless, hyaline, smooth, thin-walled, amyloid, broadly ellipsoid to ellipsoid, infrequently subglobose or elongate, infrequently reniform, usually at least somewhat adaxially flattened; apiculus sublateral, cylindric, proportionately small, infrequently rather prominent; contents mono- to multiguttulate; color in deposit not recorded. | ||||||||
| ecology |
from Yang (1997): Solitary or in small groups. China: At 1800 - 3300 m elev. In coniferous and mixed forest wtih Pinus and Picea speciesand representatives of the Fagaceae. from Tulloss et al. (2001): Solitary to gregarious, in mixed woods, mostly under conifers. Himachal Pradesh, India [Kumar et al. (1990a)]: At 2000 - 3200 m elev. In woods of Cedrus deodara, Picea smithiana and Abies pindrow or woods of C. deodara and Pinus wallichiana or under C. deodara or in dominantly coniferous forest of C. deodara and P. wallichiana with scattered Quercus incana. Pakistan: With A. pindrow. | ||||||||
| material examined |
from Yang (1997): CHINA: XIZANG AUTONOMOUS REGION (TIBET)—Linzhi Prefecture (also Nyingchi Prefecture) - Unkn. Co., unkn. loc., 3100 m elev., 19.viii.1983 Y. G. Su s.n. (HKAS 16414).
SICHUAN—Liangshan Yi Autonomous Prefecture - Xichang (county level) City, Luojishan, 2000 m elev., 16.vii.1992 M. S. Yuan 1787 (HKAS 25764, as "Amanita flavorubens" in Yuan & Sun (1995: 495)).
YUNNAN—Chuxiong Yi Autonomous Prefecture - Lufeng Co., Yipinglang, 1820 m elev., 26.vi.1978 Shengweizhan 786033 (HKAS 4530, as "A. aspera" in Zheng (1988: 120)). Dali Bai Autonomous Prefecture - Binchuan County, Jizushan, 3100 m elev. 17.viii.1985 G. P. Xiao 773 (HKAS 15417). Diqing Tibetan Autonomous Prefecture - Shangri La Co. from Yang (2002a): CHINA: JILIN—Yanbian Korean Autonomous Prefecture - Antu Co., Mt. Changbai, s.d. Y. Wang 811325 (IFP), 811038 (IFP). from Tulloss et al. (2001): INDIA: HIMACHAL PRADESH—Chamba District - Kala Top, 16.viii.1985 T. N. Lakhanpal & A. Kumar s.n. (HPUB 3324 as “A. flavoconia”). Kullu District - Manali, 11.viii.1985 T. N. Lakhanpal & A. Kumar s.n. (HPUB 3176, n.v. as “A. flavoconia”; BPI 71985 as “A. flavoconia”). Shimla District - Chopal, 29.ix.1985 T. N. Lakhanpal & A. Kumar s.n. (paratype of A. watlingii, HPUB 3619, n.v.; paratype of A. watlingii, BPI 71992). UTTARAKHAND—Pauri District - Garhwal, Nagdeva-Jhandidhar, 23.viii.1993 V. K. Bhatt & R. P. Bhatt s.n. (GUH M-20069 as “A. flavoconia”; RET 300-3). PAKISTAN: NORTHWEST FRONTIER PROV.—Hazara District - Nathiagali, Kuzagali, 7.viii.1994 A. N. Khalid 781994 (LAH; RET 140-8). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita flavipes |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita flavipes |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.