


| name | Amanita curtipes |
| name status | nomen acceptum |
| author | E.-J. Gilbert |
| images |
 1. Amanita curtipes, Larache, Morroco.  2. Amanita curtipes, Bajol, Spain. |
| intro | For the moment, please, refer to the technical tab for further information. |
| spores | The spores of this species measure (7.0-) 10.2 - 14.9 (-21) × (4.0-) 5.0 - 6.8 (-8.2) μm and are amyloid and elongate to cylindric, rarely ellipsoid or bacilliform. Clamps are not present at bases of basidia. |
| discussion |
Amanita curtipes is a species of the Mediterranean region with locations known from both southern Europe and the coastal region of North Africa. In France, it is found growing in acidic volcanic soil of mediterranean maquis and with Pines. In Morocco, it has been found in association with Pines and Oaks. The reader may wish to compare this species with A. valens, which at least until recently has been less well known.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita curtipes | ||||||||
| author | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl. (2): 279. | ||||||||
| name status | nomen acceptum | ||||||||
| synonyms |
≡Amidella curtipes E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 77, tab. 22 (figs. 3-6), tab. 23 (fig. 1).
≡Amanita baccata f. minor Bres. 1928. Iconogr. Mycol. (Milan) 6: pl. 300. [Amanita baccata misapplied by Bresadola.]
=Amanita baccata sensu Torrend. 1909. Brotéria 8: 92, pl. 3 (fig. 6).
=Amanita baccata sensu E.-J. Gilbert. 1925. Bull. Trimestriel Soc. Mycol. France 41: 287. [Misapplication.]
??=Amanita cordae Velen. 1920. České Houby 1: 191-192. [This name has also been proposed as a synonym of A. eliae Quél.]
=Amanita baccata sensu Bres. 1927. Iconogr. Mycol. (Milan) 1: 19, pl. 19.
≡Amanita baccata f. major Bres. nom. illeg. 1928. Icon. Mycol. (Milan) 6: 300. [Posterior homonym. ICBN §53.1. Intended synonym for autonym of Amanita baccata f. baccata. ICBN §11.6, §32.7.]
=Amanita valens (E.-J. Gilbert) Bertault sensu Bertault. 1980. Bull. Trimestriel Soc. Mycol. France 96(3): 281. [Misapplication.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 284052, 284130, 258045 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| lectotypes | [Note: syntypes in herb. Bresadola (S)?] | ||||||||
| selected illustrations | Romagnesi. 1961. Nouv. Atl. Champ. 3: pl. 183A. | ||||||||
| intro |
Olive text indicates a specimen that has not been
thoroughly examined (for example, for microscopic details) and marks other places in the text
where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of another researcher is based on original research by of C. Bas [notes and collection annotations in L] and R. E. Tulloss. | ||||||||
| pileus | (30-) 35 - 100 mm wide, whitish to pale pinkish buff, at times with yellowish to yellowish salmon to creamy-pale-ochraceous-pinkish tints, becoming reddish-brown over disc in age, hemispherical then convex with flattened umbo, then convex to planoconvex with depressed center; context white, sometimes pinkish under pileipellis, firm, turning pinkish when cut; margin not or very slightly striate, strongly rounded, appendiculate just at first; universal veil absent or as one or more membranous, layered patches, occasionally with paper-thin, inner layer projecting beyond some edges of thick patch formed from interior and external layers; pileipellis separable, sometimes extending beyond margin. | ||||||||
| lamellae | free to narrowly adnate with decurrent line on stipe, moderately close, rarely forking, whitish to cream with pinkish-buff reflections, drying 7E4, 5YR 5/8, 5YR 7/6, 7.5YR 6/4, 7.5YR 5-7/6, 7.5YR 7/8, 10YR 7/6-8, 8 - 14 mm broad, remarkably broad and rounded at distal end; margin flocculose, white then reddish brown in places; lamellulae truncate. | ||||||||
| stipe | 35 - 95 (-140) × 7 - 15 (-30) mm, white, discoloring like pileus, slightly flocculose to subsquamulose, upper part with minute flocculose scales turning somewhat sordid buff or pinkish buff like remainder of stipe, with base 12 - 30 mm wide; context white turning pink or very slightly pinkish-brownish when cut, stuffed, with some hollow chambers; exannulate; universal veil as rather thick-limbed, ovoid to claviform volval sac, originally whitish, exterior surface darkening/graying, interior changing color like stipe. | ||||||||
| odor/taste | Odor somewhat like a bakery (C. Bas' annotation of Bas 5534). | ||||||||
| pileipellis | 75 - 80 µm thick, lacking distinct sub- and suprapellis, gelatinizing only at surface; filamentous, undifferentiated hyphae 2.1 - 4.9 µm wide, branching, sometimes having subrefractive yellowish walls, partially gelatinized and ungelatinized, interwoven in part but dominantly subradially arranged (sometimes aggregated in radial fascicles), densely packed vertically, with terminal cells sometimes slightly inflated; vascular hyphae not observed. | ||||||||
| pileus context | filamentous, undifferentiated hyphae 4.2 - 10.5 µm wide, frequently branching, interweaving, in fascicles and singly in interstices; acrophysalides moderately densely packed, ovoid to subglobose, up to 144 × 63 µm, with walls thin or up to 1.2 µm thick; vascular hyphae not observed. | ||||||||
| lamella trama | bilateral; wcs = 20 - 30 µm (good rehydration); central stratum containing slightly inflated intercalary hyphal segments; subhymenial base with angle of divergence obscure to 20° - 40°, with elements of subhymenial tree becoming perpendicular to central stratum in subhymenium; central stratum ??; filamentous, undifferentiated hyphae 4.9 - 11.9 µm wide, branching; inflated cells as very long usually intercalary expanded hyphal segments to 25 µm wide (e.g., 91 × 25 µm); vascular hyphae 1.8 - 10.0 µm wide, sinuous, sometimes loosely coiling, scattered, absent in some sections, sometimes in clusters. | ||||||||
| subhymenium | ??; often dominated by inflated cells, but usually with branching structure clearly evident; comprising pyriform to ovoid to subglobose to clavate cells and uninflated hyphal segments in chains, in 2 - 3 layers, with basidia arising from inflated cells and (occasionally) from partially inflated or uninflated hyphal segments. | ||||||||
| basidia |
39 - 70 × 8.8 - 12.0 µm, thin-walled, predominantly 4-spored (even in relatively immature material), some 2- and 1-spored, with sterigmata up to 4.5 × 1.7 µm; clamps rare, small, most easily found in immature regions of hymenium. [Note: The presence of clamps should be rechecked.—ed.] | ||||||||
| universal veil | On stipe base, exterior surface: filamentous, undifferentiated hyphae 2.4 - 7.3 µm wide, branching, partially to rather extensively gelatinized, singly and in fascicles, sometimes dominantly in longitudinally oriented fascicles, but always with some interweaving branches, densely packed in cross-section, in layer 15 - 20 µm thick, with some hyphae just below surface sometimes having short intercalary segments with ovoid inflated (herniated) regions, sometimes below the latter zone having very thin region of sublongitudinally arranged closely-packed hyphae sometimes gelatinized and, thus, obscured?; refractive hyphae 2.0 - 5.2 µm wide, possibly product of gelatinization. On stipe base, interior: filamentous, undifferentiated hyphae 2.8 - 9.1 µm wide, branching, loosely interwoven, plentiful to dominating, in fascicles or singly, sometimes with slightly inflated intercalary segments, often constricted at septa; inflated cells at least occasionally in small clusters, locally dominating, subglobose to broadly ellipsoid to ellipsoid or ovoid to clavate, up to 90 × 74 µm, with walls thin to slightly thickened, with proportionately narrower cells more frequently thin-walled and easily damaged in sectioning, with walls of other cells occasionally up to 1.2 µm thick; vascular hyphae 1.2 - 9.0 µm wide, with occasional groups of closely spaced constrictions, rare to infrequent to locally moderately common. On stipe base, inner surface: frequently carried away (at least in part) by expanding pileus, comprising 15± µm thick layer of partially to largely gelatinized filamentous, undifferentiated hyphae dominantly longitudinally oriented; vascular hyphae not observed. On pileus, exterior surface: as on base, but more compressed. On pileus, interior: as on base except more compressed and slightly more gelatinized. On pileus, inner surface: ?dominated by interwoven frequently branching filamentous undifferentiated hyphae 1.2 - 11.9 µm wide; inflated cells scattered, clavate to subglobose to ovoid, terminal, to 123 × 82 µm, the larger (non-clavate) cells with walls up to 1.0 µm thick; vascular hyphae 4.2 - 5.6 µm. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous undifferentiated hyphae 2.1 - 9.8 µm wide, branching; acrophysalides narrow, mostly thick-walled, to 203 × 41 µm, with walls 0.5 - 1.2 µm thick; vascular hyphae 2.1 - 9.1 µm wide, infrequent. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue | not examined. | ||||||||
| basidiospores |
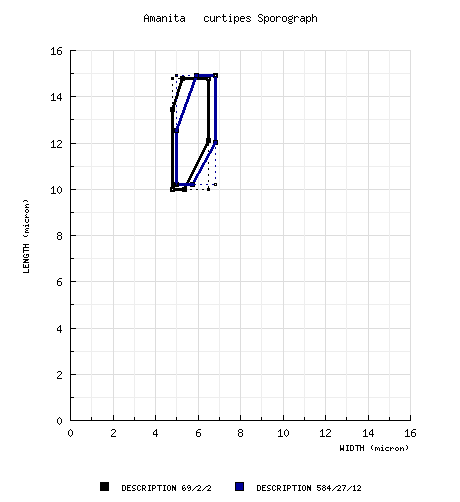
spores from topotypes (RET): [69/2/2] (9.8-) 10.0 - 14.8 (-16.0) × (4.6-) 4.8 - 6.5 μm, (L = 11.4 - 13.2 μm; L' = 12.2 μm; W = 5.3 - 5.8 μm; W' = 5.5 μm; Q = (1.69-) 1.86 - 2.80 (-3.02); Q = 2.16 - 2.29; Q' = 2.22), hyaline, colorless, smooth, thin-walled, amyloid, elongate to cylindric, infrequently bacilliform; apiculus sublateral, broadly cylindric; contents granular to guttulate; color in deposit not recorded. composite spore data from material examined by RET: [584/27/12] (7.0-) 10.2 - 14.9 (-21) × (4.0-) 5.0 - 6.8 (-8.2) μm, (L = (10.1-) 10.9 -14.0 μm; L' = 12.8 μm; W = (4.9-) 5.1 - 6.4 (-6.8) μm; W' = 5.9 μm; Q = (1.43-) 1.77 - 2.51 (-3.15); Q = (1.84-) 2.02 - 2.32 (-2.39); Q' = 2.16), hyaline, colorless, smooth, thin-walled, amyloid, elongate to cylindric, rarely ellipsoid or bacilliform, sometimes swollen at one end, often adaxially flattened; apiculus sublateral, broadly cylindric to truncate-conic; contents granular to guttulate; color in deposit not recorded. | ||||||||
| ecology | Scattered to gregarious. France: In acid volcanic soil of mediterranean maquis including Pinus sylvestris. Morocco: Under Pinus pinea or with Quercus suber. | ||||||||
| material examined | FRANCE: ALPES MARITIMES—Berre-des-Alpes, vi.1910 G. F. Atkinson 24591 (topotype, FH), 20.vi.1926 E.-J. Gilbert s.n. (topotype, L). LANDES—St-Martin-d'Oney, 17.xi.1965 J. Beller & M. Bon 2937 (MPU), Yrieu, v.1954 A. G. Parrot 7505 (L as "A. gilbertii f. minor" [herb. name—ed.]). VAR—Massif de l'Estérel, 25.iv.1971 J. Trimbach s.n. (L). ITALY: SARDINIA—Costa Paradiso, 3.xi.1983 M. Moser 83/539 (IB as "A. gilbertii"). MOROCCO: Donabo, 1.xi.1972 R. Bertault 12315 (L). El Araïche (Larache), bois de Larache, 11.i.1966 R. Bertault 10790 (L as "A. valens sensu Bertault"), R. Bertault 10791 (L as "A. valens sensu Bertault"), 3.ix.1967 R. Bertault 11071 (L as "A. valens sensu Bertault"), 22.iv.1971 R. Bertault & C. Bas 5534 (L). Khermiss-de-Sahel, 3.iii.1972 R. Bertault 12293 (L). | ||||||||
| discussion |
Bresadola (1928?) illustrates cylindric spores and a cellular subhymenium for his A. baccata f. minor which is in good agreement with our interpretation of A. curtipes; he separated f. minor from A. baccata f. major (an inadmissible name intended to be equivalent to A. baccata f. baccata) by its slightly smaller size and by the fact that its pileus diameter was nearly the same as the length of its stipe (in comparison to the more slender habit of “f. major”). Amanita baccata f. major is also illustrated; and, again, cylindrical spores with size in the range expected for A. curtipes and a cellular subhymenium are shown. While no holotype is specified, the locality given is “Italia centralis (Latium).” Central Italian material from Bresadola’s herbarium should be selected as lectotype for Bresadola's f. minor. Gilbert (1930?) recognized that Bresadola had been mistaken and that the name baccata should be applied to a member of section Lepidella. Later, he created the epithet curtipes for A. baccata f. minor. Gilbert’s collection on which he based his description of A. curtipes is apparently lost along with all of Gilbert’s personal herbarium. The collection would be dated 20.ix.1925, made by E. J. Gilbert; and, at one time, there was a watercolor of the collection. Apparently, the watercolor is also lost. Amanita curtipes is usually thought of as a semi-russuloid, small species with the stipe usually shorter than the pileus diameter. This opinion enshrined in the specific epithet has led to some confusion. The most recent difficulty arose when Bertault (1980), while correcting an oversight of Kühner & Romagnesi in neglecting to cite a basionym in making the combination A. valens (E.-J. Gilbert) Bertault, misapplied the name to collections of A. curtipes which he segregated from others of the same species based on their larger habit. However, he had difficulties with his concepts of the two species. Bertault (1980) records an incident in which he was shown two collections of A. curtipes that apparently had been growing from the same mycelium, but was not told about the conditions of their collection. He unhesitatingly separated them as A. curtipes and A. valens; only to find himself in error when all the information about the collections was made known to him. While he states he learned much from the experience, he continued to maintain size range of the basidiocarps as the determining character for A. curtipes and continued to apply the name A. valens to large specimens of A. curtipes. While the concept of A. valens may still not be completely grasped today, the entity to which the name applies certainly must have ellipsoid to elongate rather than cylindric spores (Gilbert, 1941). Most modern authors, other than Bertault, agree that A. valens has ellipsoid spores as a consequence of their being considerably broader than the spores found in A. curtipes. The following diagram compares sporographs of the two species: Notice that using spore size and shape to segregate the taxa seems to work rather well. For comparison we provide here the spore measurement statistics from the European mainland, Sardinian, and African collections of A. curtipes which we examined: BASIDIOSPORES from African collections: [375/17/6] (7.0-) 10.1 - 15.0 (-17.2) × (4.9-) 5.5 - 6.8 (-8.2) µm, (L = (10.1-) 11.9 - 14.0 µm; L’ = 13.0 µm; W = (5.6-) 5.8 - 6.4 (-6.8) µm; W’ = 6.0 µm; Q = (1.43-) 1.77 - 2.50 (-3.15); Q = (1.81-) 2.04 - 2.32 (-2.39); Q’ = 2.16).Amanita ponderosa Malenç. & Heim apud Malenç. can be easily separated from A. curtipes by the often massive habit of the former, its subhymenium which consists of cylindrical hyphae perpendicular to the central stratum of the trama, and its broader spores. Amanita lepiotoides Barla frequently has thick scales on its pileus representing the interior layer(s) of its universal veil which forms a large, elongate sac at the stipe base; and the subhymenium of A. lepiotoides is rather like that of A. ponderosa (definitely not pseudoparenchymatous). Amyloid warts were relatively common on the spores of Bertault 12315. In cross-section they appear as a slight, external, thickened area of the spore wall. The portion of Parrot 7505 in L is much impregnated with sand and probably was attacked by a mold rather extensively before or after drying. While many characters are obscured, the spores, subhymenium, and universal veil were sufficiently similar to the Moroccan collections of Bertault and Bas so that we place the collection here. However, the universal veil tissue was considerably collapsed; and, probably as a consequence, the hyphae in the interior layer showed more tendency to a longitudinal orientation than was seen in the African material. Moser and Jülich (1985-1995) illustrate Moser 83/539 with a color photograph. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita curtipes |
| bottom links | [ Keys & Checklists ] |
| name | Amanita curtipes |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.