


| name | Amanita constricta |
| name status | nomen acceptum |
| author | Thiers & Ammirati |
| english name | "Great Gray-Sack Ringless Amanita" |
| images |
 1. Amanita constricta, northern California, U.S.A.  2. Amanita constricta, mature graying volval patch on stipe, northern California, U.S.A. .jpg "Amanita constricta, mature graying volval, Del Norte Co., California, U.S.A. (RET 817-8)")  3. Amanita constricta, mature graying volval, Del Norte Co., California, U.S.A. (RET 817-8) ") 4. Amanita constricta, mature graying volval, Del Norte Co., California, U.S.A. (RET 817-8) |
| intro | The information below is based on a type study and other original research of RET. |
| cap | The cap of A. constricta is 50 - 130 mm wide, convex when young, becoming plano-convex to plane, eventually subumbonate to umbonate in age, with a strongly sulcate to tuberculate striate margin (20 - 25% of the radius). The cap is brown to brownish gray, the disc at times is as dark as Chaetura black, often with inconspicuous dark radial streaks. The flesh is white, sometimes becoming faintly pinkish with exposure (see discussion below). The volva is absent or present as a membranous fibrillose patch over the cap's center; it is white to buff to (eventually) smoke gray. Although I have never seen it, the volva is said to bruise salmon color when moistened (see discussion below). |
| gills |
The gills are close to crowded, adnate to decurrent by a short hook when young, becoming free, white at first, becoming gray, and drying tan to sordid tan to brownish gray. The gills are moderately broad with the edge usually gray and fibrillose. The short gills are in several ranks. |
| stem | The stem is 100 - 160 (-200) mm long, 7- 17 mm wide at the apex, white, cylindric or narrowing upward, and exannulate. The flesh is white, stuffed, becoming hollow. The membranous volva soon begins to turn gray and become structurally weaker; although I have never observed this, it is said to bruise reddish or salmon when wet then fade back to its previous coloration (see discussion below). |
| spores | The spores measure (7.2-) 9.5 - 12.8 (-19.0) × (6.2-) 8.2 - 11.5 (-17.8) µm and are inamyloid and globose to subglobose to broadly ellipsoid (occasionally ellipsoid). Clamps are probably absent from bases of basidia. |
| discussion |
Associated with Pacific coastal species of oak and arbutus as well as Douglas Fir. Described originally from California, this species range probably extends into southwestern Canada. The original description was based on collections of several different species including A. protecta Tulloss & G. Wright and undescribed taxa. Since there is distinct rusty or ochraceous staining in all parts of A. protecta, this could be the source of the claim that the volva of A. constricta stains. The original description's statement that the cap of the present species can be gray may also result from inclusion of A. protecta in the original concept of A. constricta. At present, no taxa are known to me that appear particularly closely related to A. constricta. The reader may wish to compare the taxa similar to A. submembranacea (Bon) Gröger.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita constricta | ||||||||||||||||||||||||||||||||||||||||
| author | Thiers & Ammirati. 1982. Mycotaxon 15: 163. | ||||||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||||||
| english name | "Great Gray-Sack Ringless Amanita" | ||||||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 261724 | ||||||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||||||
| holotypes | SFSU | ||||||||||||||||||||||||||||||||||||||||
| type studies | Tulloss. 1994. Mycotaxon 52: 321. | ||||||||||||||||||||||||||||||||||||||||
| revisions | Tulloss, here | ||||||||||||||||||||||||||||||||||||||||
| intro |
The following text may makes multiple use of each data field. The field may contain magenta text representing data specifically from a type study by Tulloss. The same field may also contain black text, which will represent a revision of the species by Tulloss. Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following is based on the field notes with the holo- and paratypes, on the text of the protolog, and on original research of R. E. Tulloss. | ||||||||||||||||||||||||||||||||||||||||
| pileus | Tulloss (1994): 50 - 130 mm wide, gray to brownish gray, Pale Mouse Gray to Light Drab Gray to Drab to Hair Brown, disc at time as dark as Chaetura Black, often with inconspicuous dark radial streaks, convex when young, becoming plano-convex to plane, eventually subumbonate to umbonate in age; context white except next to the pileipellis where it is gray to drab, sometimes become faintly pinkish on exposure especially in young basidiomes, 5 - 17 mm thick at disc, thinning toward margin; margin decurved at first, soon plane to undulate, slightly uplifted in age, en-tirely becoming rimose, strongly sulcate to tuberculate striate (0.2 - 0.25R); universal veil absent or as a membran-ous somewhat fibrillose patch over disc, typically less than 1 mm thick, sometimes breaking up around the edges in age, white to buff to Smoke Gray or sometimes with pale vinaceous shades, bruising salmon color when wetted, slowly returning to white. | ||||||||||||||||||||||||||||||||||||||||
| lamellae | Tulloss (1994): close to crowded, adnate to decurrent by a short hook when young, becoming free, white at first becoming gray, Smoke Gray to Pallid Mouse Gray to Pale Drab Gray, drying tan to sordid tan to brownish gray, (grayer than 7.5YR 6/6 to 10YR 5/4 to 10YR 7-8/4), moderately broad; edge usually gray, fimbriate; lamellulae in several ranks. | ||||||||||||||||||||||||||||||||||||||||
| stipe | Tulloss (1994): 100 - 160 (-200) mm long, 7 - 17 mm wide at apex, white, cylindric or narrowing upward, apex longitudinally striate or pruinose, becoming appressed fibrillose below, stipipellis often rupturing and forming rings of appressed fibrillose scales which are gray and darken when bruised; context white, stuffed, becoming hollow; exannulate; universal veil membranous, white to pale buff or pale yellow, bruising reddish when wet then fading, becoming gray and submembranous at maturity and then rather easily fragmented and sometimes remaining in large patches in substrate or on lower stipe, adnate to stipe for one-third to one-half of limb length, then flaring in a manner suggesting an annulus, collapsing with age. | ||||||||||||||||||||||||||||||||||||||||
| odor/taste | Tulloss (1994): Odor and taste not distinctive. | ||||||||||||||||||||||||||||||||||||||||
| macrochemical tests |
Breckon (1968)—on exposed context of the lower stipe and bulb unless otherwise noted: Concentrated H2SO4 - turning lamellae pink to lilac, but not always; FeSO4, 15% KOH, Melzer’s Reagent - negative; 3% phenol - positive; tincture of guaiac - varying from strongly and rapidly positive to weakly and slowly positive; phenolaniline - rapidly positive. Any variable reactions must be questioned because Breckon had as many as four taxa mixed together among the collections cited under the present species in his thesis. | ||||||||||||||||||||||||||||||||||||||||
| pileipellis | Tulloss (1994): 200± µm thick; thin layer at surface gelatinized; upper 40± µm incompletely gelatinized; upper 80 - 95 µm hyaline to pale yellowish; lower portion light brownish orange (mounted in 10% NH4OH); filamentous, undifferentiated hyphae 2.2 - 8.5 µm wide, criss-crossed, largely uncurving, many subradially arranged, a few containing red-brown granules (mounted in 3% KOH); vascular hyphae not observed. | ||||||||||||||||||||||||||||||||||||||||
| pileus context | Tulloss (1994): filamentous, undifferentiated hyphae 1.5 - 14.0 µm wide, branching, very loosely interwoven; acrophysalides dominating, subglobose to pyriform to ellipsoid to broadly clavate to clavate, up to 129 × 74 µm; vascular hyphae 6.0 - 17.5 µm wide, loosely coiling. | ||||||||||||||||||||||||||||||||||||||||
| lamella trama | Tulloss (1994): bilateral, with divergence obscured by plentiful hyphae parallel to the hymenial surface and the tangled nature of the subhymenial tree except near the edge of the lamella; central stratum having boundaries somewhat difficult to define because of density of subhymenium and fact that divergence begins within central stratum and many hyphae of the subhymenial base diverge at very shall angles which are maintained for relatively long distances, sometimes imperfectly rehydrating and then dense and brown to orange-brown, with wcs = 110 - 145 µm in cases of most complete rehydration (65 - 85 µm in most common degree of moderate rehydration, 30 - 55 µm in poorly rehydrating trama); with hyphae of subhymenial base diverging at angles from very shallow to over 60°; filamentous, undifferentiated hyphae 1.8 - 9.8 µm wide, sometimes coiling, interwoven, frequently branching, with central stratum and subhymenial base including intercalary inflated cells [thin-walled, subventricose to ventricose to subclavate to clavate to broadly clavate to irregular, up to 53 × 35 µm (most often about half this width)]; divergent, terminal, inflated cells not observed; vascular hyphae 2.5 - 12.5 µm wide, occasionally branching, infrequent to somewhat common to common. | ||||||||||||||||||||||||||||||||||||||||
| subhymenium | Tulloss (1994): wst-near = 80 - 140 µm in cases of most complete rehydration [(25-) 30 - 80 µm in most common degree of moderate rehydration, 0 - 30 µm in poorly rehydrating trama]; wst-far = 110 - 175 µm in cases of most complete rehydration (60 - 105 µm in most common degree of moderate rehydration, 35 - 60 µm in poorly rehydrating trama); a complexly interwoven structure comprising frequently septate, frequently branching hyphae, with segments uninflated or partially inflated (or occasionally inflated and then clavate to subpyriform to somewhat irregular, e.g., 19 × 14 µm), with hyphae of the subhymenium occasionally running parallel to the hymenial surface (sometimes in rather dense clusters) within 10 µm of bases of longest basidia, with basidia often arising from uninflated segments (80% or more of cases examined), but also from any type of cell cited, with 1/2 to 2 1/2 (rarely 3) cells between base of short basidiole and longest nearby basidium/ ole, with cell immediately below some short basidioles uninflated (9.0 - 21 × (2.5 ) 3.2 - 6.8 µm) to narrowly clavate or with short branch giving rise to second basidiole (12.6 - 20 × 5.8 - 8.8 µm) to ellipsoid or pyriform (7.5 - 10.0 × 7.8 - 11.8 µm). | ||||||||||||||||||||||||||||||||||||||||
| basidia |
Tulloss (1994):
54 - 78 × 8.5 - 19.5 µm,
thin-walled, dominantly 4-, but occasionally 1- or
2-sterigmate; sterigmata up to 16.0 × 3.5 µm; clamps
rather rare to rather common. [This report of clamps is unlikely to be correct.—ed.] | ||||||||||||||||||||||||||||||||||||||||
| universal veil | Tulloss (1994): On pileus, upper portion: filamentous, undifferentiated hyphae 1.5 - 8.2 µm wide, occasionally in fascicles, often collapsed; inflated cells dominating, thin-walled, clavate to broadly clavate to subpyriform to ellipsoid, often pale brownish or pale orange-brown, terminal, single, up to 46 × 33 µm, collapsed; vascular hyphae not observed. On pileus, lower portion (adjacent to pileipellis): hyphae more frequent; all elements extensively compressed. On stipe about 10 mm above base: structures similar to those on pileus; inflated cells (collapsed and partially gelatinized) and fragments of same easily dissociated from surface leaving mostly tangled hyphae; inflated cells up to 95 × 67 µm, with walls thin or up to 0.8 µm thick, with unbroken cells often not pigmented; vascular hyphae 3.0 - 6.0 µm, scant, fragmentary. | ||||||||||||||||||||||||||||||||||||||||
| stipe context | Tulloss (1994): longitudinally acrophysalidic; filamentous, undifferentiated hyphae 1.5 - 11.5 µm wide, branching, plentiful; acrophysalides thin-walled, plentiful, up to 271 × 45 µm, those of broader diameter often broadening rapidly close to basal septum; vascular hyphae 2.8 - 10.5 µm wide, loosely coiling. | ||||||||||||||||||||||||||||||||||||||||
| lamella edge tissue | not described | ||||||||||||||||||||||||||||||||||||||||
| basidiospores |
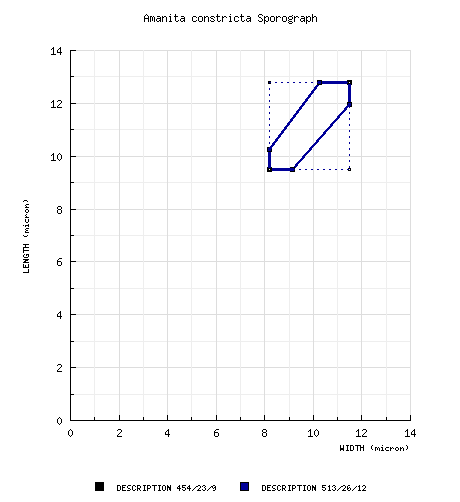
Tulloss (1994): [454/23/9] (7.2-) 9.5 - 12.8 (-19.0) × (6.2-) 8.2 - 11.5 (-17.8) μm, (L = (10.3-) 10.4 - 11.8 μm; L' = 11.0 μm; W = (8.8-) 9.0 - 10.5 (-10.9) μm; W' = 9.9 μm; Q = (1.0-) 1.04 - 1.25 (-1.56); Q = (1.06-) 1.07 - 1.17 (-1.18); Q' = 1.12). composite of data from all material revised by RET: [513/26/12] (7.2-) 9.5 - 12.8 (-19.0) × (6.2-) 8.2 - 11.5 (-17.8) µm, (L = (10.3-) 10.4 - 11.7 (-11.8) µm; L’ = 11.0 µm; W = (8.8-) 9.2 - 10.5 (-10.9) µm; W’ = 9.9 µm; Q = (1.0-) 1.04 - 1.25 (-1.56); Q = (1.06-) 1.09 - 1.17 (-1.18); Q’ = 1.12), colorless, hyaline, smooth, thin-walled, inamyloid, globose to subglobose to broadly ellipsoid, occasionally ellipsoid, in some specimens frequently lacrymoid, adaxially flattened (often strongly so), often swollen at one end, suceptible to conversion to crassospores on occasion (Tulloss 1-19-03-Ba); apiculus sublateral, cylindric to truncate-conic, at times quite prominent; contents mono- to multiguttulate to granular; white in deposit. | ||||||||||||||||||||||||||||||||||||||||
| ecology |
from type study of
(1994):
Single to scattered to
gregarious. Along the Pacific Coast of the
U.S.A., apparently limited to coastal forest, under
hardwoods such as coastal spp. of Quercus
(e.g., Q. agrifolia) and Arbutus
menziesii, on occasion under Pseudotsuga
menziesii or in mixed woods of P.
menziesii and Q. agrifolia or under
Umbellularia californica in an area of
mixed Quercus and
Umbellularia. The holotype was
collected under Quercus and A.
menziesii. California: In wet soil with Picea, Pinus, and Lithocarpus densiflorus (Tanoak) or in forest dominated by Picea sitchensis. | ||||||||||||||||||||||||||||||||||||||||
| material examined | RET: U.S.A.: CALIFORNIA—Del Norte Co. - Jedediah Smith St. Pk., Howland Hill Tr. [41.7844° N/ 124.1035° W, 213 m], 3.i.2018 Ronald L. Pastorino 1-3-18B [mushroomobserver #305220] (RET 817-8, nrITS seq'd.). Humboldt Co. - Prairie Creek Redwoods St. Pk. [1.4154° N/ 124.0455° W, 146 m], 15.i.2014 Hunter Sezen s.n. [mushroomobserver #162877] (RET 666-9, nrITS & nrLSU seq'd.). Marin Co. - Alpine Lk., 20.xii.1963 Harry D. Thiers 11146 (paratype, SFSU); Muir Woods Nat. Mon., 1.i.1967 D. E. Madden 904 (paratype, SFSU), 25.i.1967 Gary A. Breckon 881 (paratype, SFSU), 31.i.1967 G. A. Breckon 906 (paratype, SFSU). Mendocino Co. - W of Navarro, Cameron Rd., 19.i.2003 Howard Higley s.n. [Tulloss 1-19-03-Ba] (RET 366-6, nrITS seq'd.). San Mateo Co. - San Francisco Watershed, 22.xii.1963 H. D. Thiers 11184 (paratype, SFSU), 11.i.1965 H. D. Thiers 12064 (paratype, SFSU), 22.i.1965 G. A. Breckon 302 (holotype, SFSU; isotype, NY 00618736), 22.ii.1965 H. D. Thiers 12211 (paratype, SFSU), 6.i.1967 G. A. Breckon & H. D. Thiers [Breckon] 674 (paratype, SFSU). Solano Co. - Green Valley, 1.iii.1968 Walter J. Sundberg 1167 (paratype, SFSU). Unkn. Co. - Redwoods Nat. Pk., Davison Rd., 20.xii.2011 Noah Siegel s.n. (RET 601-1, nrITS-LSU seq'd.). OREGON—Clackamas Co. - unkn. loc., Riverside Tr., 20.v.1987 Wanda Caruthers s.n. [Tulloss 5-20-87-WC1] (RET 244-9), 24.v.1987 W. Caruthers s.n. [Tulloss 5-20-85-WC1] (RET 244-8). WASHINGTON—Pacific Co. - Ilwaco, 11.xii.2014 Mikeal No-Line s.n. [mushroomobserver #193286] (RET 667-8, nrITS seq'd.). Whatcom Co. - Bellingham, Stimpson Nature Preserve, 21.x.2004 Buck McAdoo pg. 358#1 (RET 662-1, nrITS-LSUJ seq'd.), 27.x.2010 Buck McAdoo 212 (RET 662-7, nrITS-LSUJ seq'd.), 26.x.2013 Jack Waytz s.n. (RET 615-8, nrITS seq'd.), 27.x.2013 J. Waytz s.n. [McAdoo 12] (RET 615-9, nrITS seq'd.). | ||||||||||||||||||||||||||||||||||||||||
| discussion |
Large lacrymoid spores (not uncommon in some
specimens) are extremely difficult to measure
accurately because the large end tends to sink
producing a foreshortening of the spore as viewed,
preventing a lateral view, and making the process of
spore measurement for this taxon extremely
tedious. Lacrimoid spores were not measured
unless they could be viewed fully in focus in optical
cross-section of a lateral view. This
methodological decision tends to produce a lower than
actual Q for specimens in which lacrimoid
spores occur with any frequency. In the
specimens with lacrimoid spores, subglobose to
globose spores were always present and in equal or
greater number than the lacrimoid ones. As a
consequence, the distribution of Q for spores
measured from these specimens is often distinctly
bimodal.
| ||||||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita constricta |
| bottom links |
[ Keys & Checklists ] [ Pacific coastal states (USA) & region key ] |
| name | Amanita constricta |
| bottom links |
[ Keys & Checklists ] [ Pacific coastal states (USA) & region key ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.